
Fluid, Electrolyte, and pH Homeostasis
Blood volume, pressure, osmolarity, and pH are intimately linked, but are regulated in exquisitely complex and different ways. Sensors and effectors exist across the body, but the kidney plays a fundamental role in regulating each.
- fluid homeostasis
- electrolyte homeostasis
- pH homeostasis
Fluid Homeostasis
The body senses fluid osmolarity and volume, with the aim of maintaining blood pressure and osmolarity between 280-290 mOsm/L.
Approximately 1/5 of the cardiac output perfuses the kidney - the highest blood flow rate of any organ of the body. The glomerulus filters 25% of blood flowing through it to form ultrafiltrate due to arteriolar pressure, high permeability, and large surface area of the capillary bed.
Normal kidneys are able to autoregulate renal blood flow, keeping it constant over a range of arterial pressures. This is accomplished by modulating afferent and efferent arteriolar resistance, as well as filtration surface area by mesangial cells, via autoregulation and tubuloglomerular feedback. Autoregulation can be intefered with by ACE inhibitors and decreases with age.
Osmolarity sensors
Osmoreceptors are found in the hypothalamus and the duodenum. These receptors are activated above 290 mOsm/L, causing thirst and leading to release of anti-diuretic hormone (ADH) from the posterior pituitary. ADH acts on the connecting tubule and collecting duct to increase water resorption, concentrating the urine and increasing blood volume and pressure.
Volume sensors
Blood volume is regulated by a number of systems. The renin-angiotensin-aldosterone system (RAAS) is activated in hypovolemic states induces vasoconstriction and sodium retention.
Atrial natriuretic peptide is released by the atria in response to stretch, preventing sodium uptake and inhibiting the action of renin and aldosterone. Carotid stretch receptors also play a role in regulating blood volume.
Fluid and the Kidney
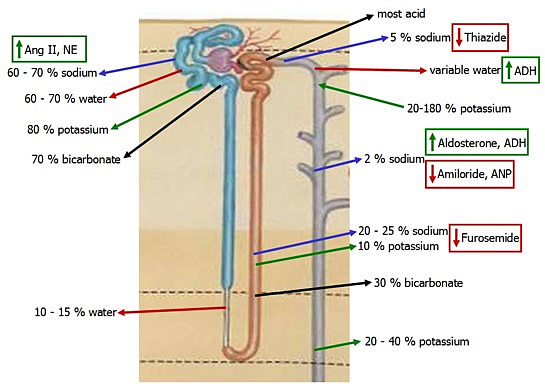
The proximal tubule absorbs 2/3 of filtered water through both aquaporin AQP1 and paracellular means. Its function is relatively invariable, compared with the distal nephron.
Tubular sodium reabsorption, mediated by the Na/H antiporter, reduces the osmolarity of the fluid, thus promoting iso-osmotic reabsorption.
The thin descending loop reabsorbs 10-15% via the hyperosmotic medullary interstitium and the presence of AQP1 in apical and basolateral membranes.
Sodium and water reabsorption are uncoupled in the Loop of Henle. Sodium reabsorption in the ascending limb creates a hyperosmotic gradient for water reabsorption in the descending limb. Only a small gradient is required to move water.
Following reabsorption from the tubule lumen, water returns to the circulation through the vasa recta. Elevated oncotic pressure in the capillaries causes absoption of both water and electrolytes from the interstitium.
The ascending LoH cannot absorb water. Reabsorption of Na/Cl/K in the thick ascending limb via the very effecieint triportercreates hypoosmotic fluid 1/3 that of plasma, as the distal tubule and collecting duct are normally water impermeable.
The countercurrent multiplier in the loop of Henle establishes a gradient from 300 mOsm/kg H2O to 1200 mOsm/kg H2O, extending from the corticomedullary junction to the tip of the papilla. This gradient drives water from the thin descending limb and from the medullary collecting duct in the presence of ADH.
The osmotic gradient is at most 200 mOsm/kg H2O at any point, as established by both sodium and urea.
Urea deposition contributes to half the osmolarity of the medullary interstitium, facilitated by ADH-mediated urea transporter activity. This helps maintain the NaCl gradient in the medullary interstitium. However, urea does not draw water out of the collecting duct, as it is membrane-permeable and does not establish a gradient.
If the countercurrent multiplier were based on sodium alone, the max osmotic gradient would be 600.
Slow medullary blood flow, as well as the arrangement of capillaries in long hairpin loops (vasa recta), minimizes solute loss from the medullary interstitium.
Countercurrent multiplier animation
Hypovolemia
Hypovolemia is sensed by atrial baroreceptors, which communicate to the CNS. This activates the SNS and its effects on the CV system, increasing cardiac output by
- increasing heart rate and contractility
- increase venous return through increased venous tone
The long term response to hypovolemia, and to increased plasma osmolarity, is to increase plasma levels of hormones targeting the kidney:

- ADH
- norepinephrine
- angiotensin II
- aldosterone
This decreases GFR and increases sodium and water reabsorption.
Hypervolemia
change in hormone levels during hypervolemia are opposite those of hypovolemia:
- decreased ADH, Ang II, norepinephrine, and aldosterone, and
- increased ANP
This increases water and sodium excretion.
Electrolyte Homeostasis
As fluid levels are tightly linked with solute concentrations, electrolyte concentrations are of key importance.
Sodium is the most important extracellular cation and has much control on fluid levels.
Potassium levels are low extracellularly, but its fundamental importance in neuronal and cardiac function make kidney regulation highly sophisticated.
Chloride is an important extracellular anion, and it largely follows sodium.
Bicarbonate is another key extracellular anion, and acts as a pH buffer as well.
Thick Ascending Limb
While the Na/K/Cl2 triporter pumps each of these ions out, the potassium can re-enter the tubule down its concentration gradient through a ROMK channel. This leaves the lumen positively charged and can drive paracellular exit of sodium, potassium, calcium, magnesium, and ammonium.
Collecting Tubules and Ducts
Principal cells .. Sodium enters principal cells through ENaC channels, leaving a negative charge in the lumen and driving paracellular reabsorption of chloride.
Intercalated cells have no Na/K ATPase but do have an H+ ATPase, establishing a H+ concentration gradient used for transport. The net effect is secretion of bicarbonate and reabsorption of chloride.
pH Homeostasis
The kidney is an important regulator of pH due to its ability to regulate secretion and reabsorption of bicarbonate and acid.
Normally all of the bicarbonate that is filtered is reabsorbed - 70% in the proximal tubule and the rest in the loop of Henle.
Acid is secreted primarily in the intercalated cells of the distal tubule, buffered by ammonia.
The kidney responds to perturbations in pH by regulating levels of bicarbonate and acid.
Bicarbonate reabsorption is controlled by activity of the apical Na/H exchanger and basal Na/HCO3 cotransporter.
Acid secretion is controlled in the proximal tubule, as is NH3 production in the proximal tubule via breakdown of glutamate.
During periods of acidosis, the kidney secretes 1 molecule of NH4+ into the urine while adding one HCO3- back to the blood.
Most metabolic processes end with the production of acid, the largest the CO2 produced by glycolysis. The body copes with the perhaps 20,000 mEq of H+ produced daily through buffers, ventilation, and renal regulation.
The normal concentration of hydrogen in arterial blood is 40 mEq/L, yielding a pH of 7.38-7.42. Maintaining a proper pH is important to ensure intracellular proteins maintain their three-dimensional shape.
Buffers
This concentration is maintained via circulating and intracellular buffers. Their capacity is limited, however, and effective mechanisms for excretion are critical.
There is about 600,000 times as much bicarbonate than hydrogen in the blood, principally due to hemoglobin's buffering capacity.
Ventilation
Ventilation can be used to change pH by varing the amount of carbon dioxide exhaled. Peripheral cehemoreceptors in the carotid and aortic bodies sense pH, while central chemoreceptors in the medulla respond to levels of CO2.
Kidneys
The kidneys acts to excrete or resorb H+ while also changing resorption or excretion of bicarbonate.
The kidney reclaims 4500 mEq of bicarbonate daily, as well as generates new bicarbonate.
Proximal tubule
During acidosis, the kidney excretes H+ and NH4+. H+ ions comibine with phosphate to sequester it in the lumen. At the same time, the kidneys generate bicarbonate from intracellular carbonic anhydrase.
The principal process of bicarbonate generation is through urinary acidification and ammonia generation. Ammonia is generated from glutamine and secreted in the proximal tubule, which is the primary site of bicarbonate resorption. Carbonic anhydrase in the lumenal membrane rapidly converts carbonic acid to carbon dioxide and water, preventing the
Distal tubule
The distal tubule and two types of intercalated cells play an important role in fine tuning the acid-base balance. The first type secretes acid while the second secretes base, depending on polarity of the transoprt proteins. In A-type cells, HCO3- is resorbed and released basolaterally via Cl- exchange. Final titration of urinary buffers such as NH4+ is achieved by H+ secretion. In B-type intercalated cells, this process is reversed.
These cells have high levels of carbonic anhydrase intracellularly. H+ is pumped out via a H+/K+ exchanger, while HCO3- leaves via a Cl- exchanger. Acidosis is managed by type A cells, while alkalosis is managed by type B cells.
H+ Secretion
70-100 mmoles of H+ are produced and excreted daily, occurring primarily in the intercalated cells of the distal tubule.
Acid excretion normally increases linearly with acid production, which varies according to diet. Vegetarians have a more alkalotic pH than do people who eat a lot of meat, which contains many acidic amino acids.
H+ secretion by the distal nephron is limited by low pH in the urine. Phosphate and creatinine provide minimal contribution to urine's buffering capacity.
Acid is excreted in three forms:
- Free protons can be excreted to a maximum of 40 umol/l, producing a pH of 4.5. THis is as low as it can go.
- H+ can be bound to phosphate in the urine, producing titratable acidity. Phosphate can hold 10-40 mmol/day.
- The majority of H+ are excreted in ammonium, with 30-40 mmol/day excreted. This can increase 10x. Ammonia can diffuse freely, but charged ammonium cannot, a phenomenon called diffusion trapping.

As H+ is being pumped out of the cell against its concentration gradient, both secretion mechanisms require energy.
Defects in the proton pump, in ammonia production, or H+ backleak can cause metabolic acidosis.
Ammonia Metabolism
Ammonia is a great buffer and works by binding H+ and allowing maximal H+ secretion into the urine. The kidney produces 30-300 mmoles of NH4+ daily for excretion in the urine.

Ammonia is what gives urine its smell, and more of it is produced following protein digestion.
Ammonia is secreted in the proximal tubule as a component of bicarbonate reabsorption following production of alpha-ketoglutarate from glutamate. This alpha-ketoglutarate is used in gluconeogenesis.
Ammonia accumulates in the medullary interstitium, and plays a role in carrying H+ in the urine of the distal tubule as ammonium.
Acidemia increases the ratio of NH3 to NH4+, while alkalemia reduces it.